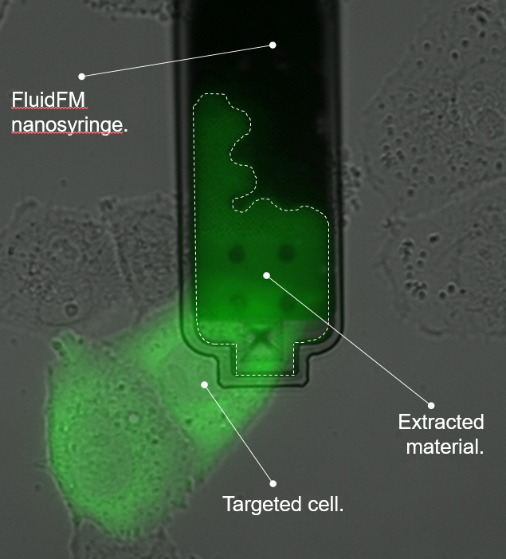
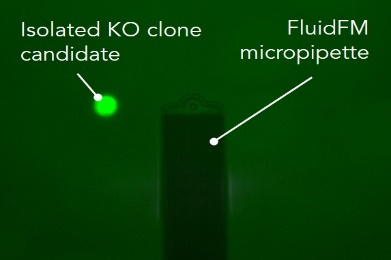
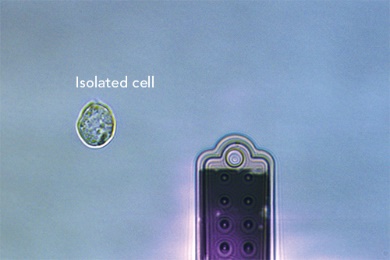
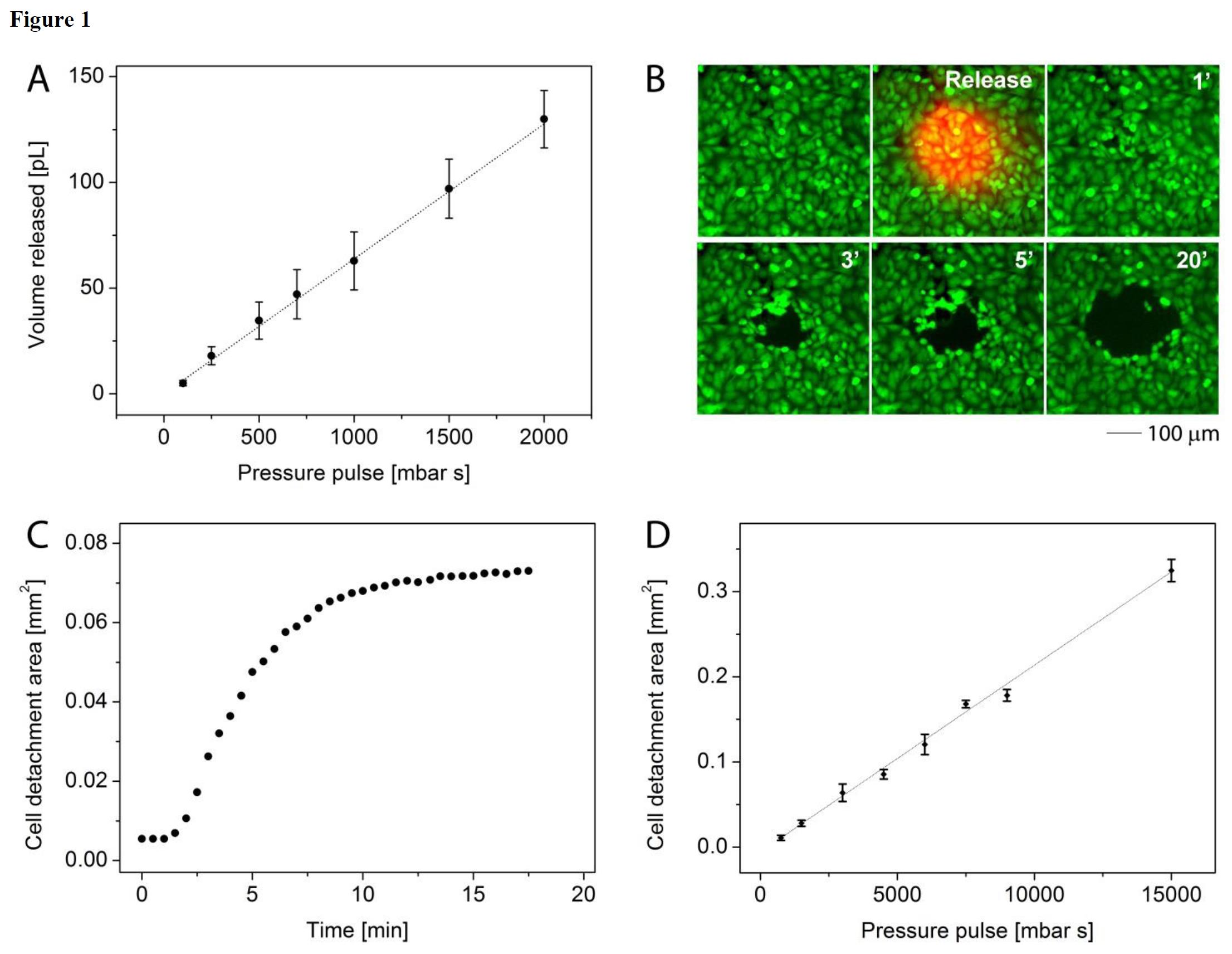
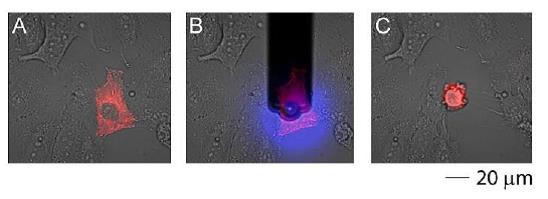
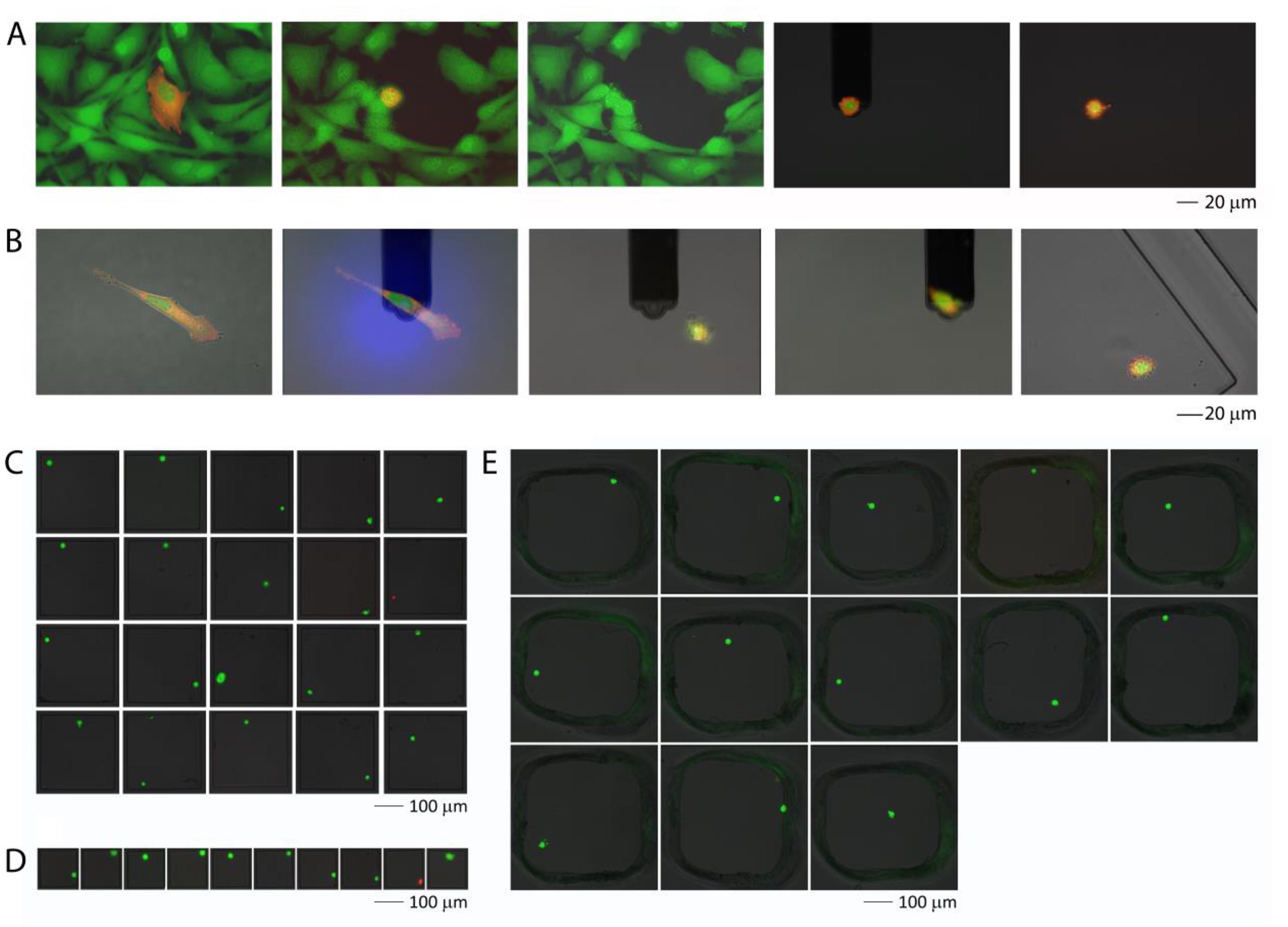
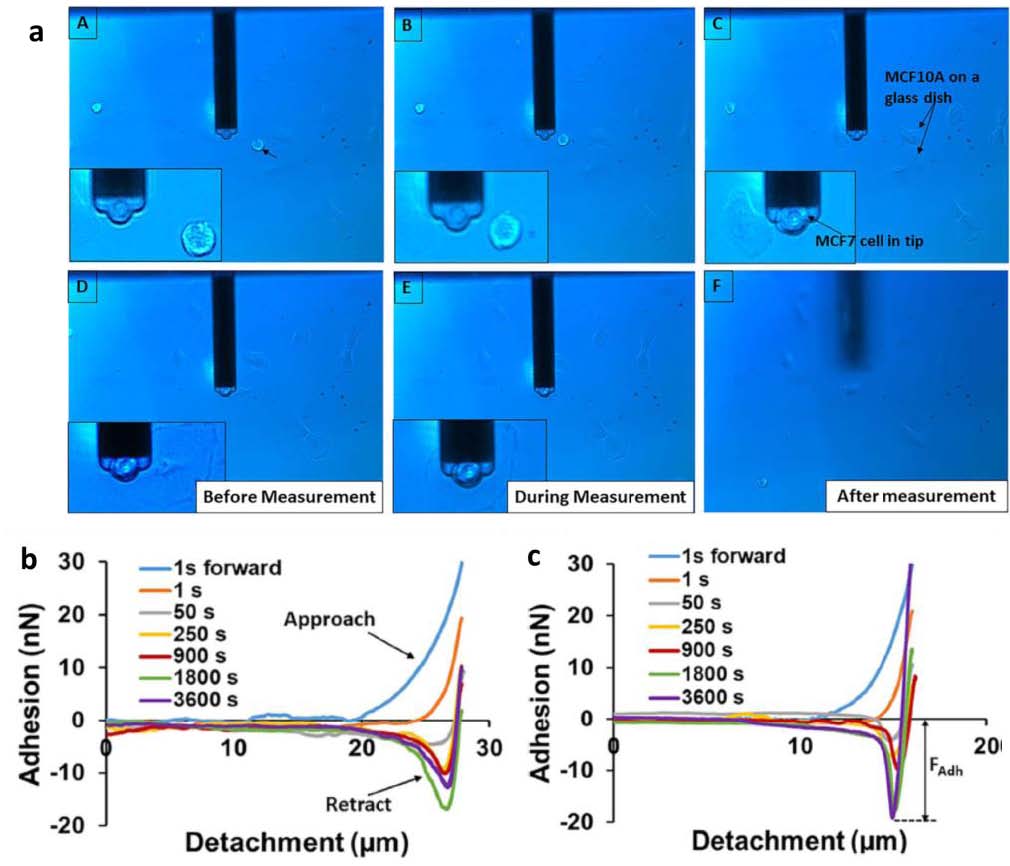
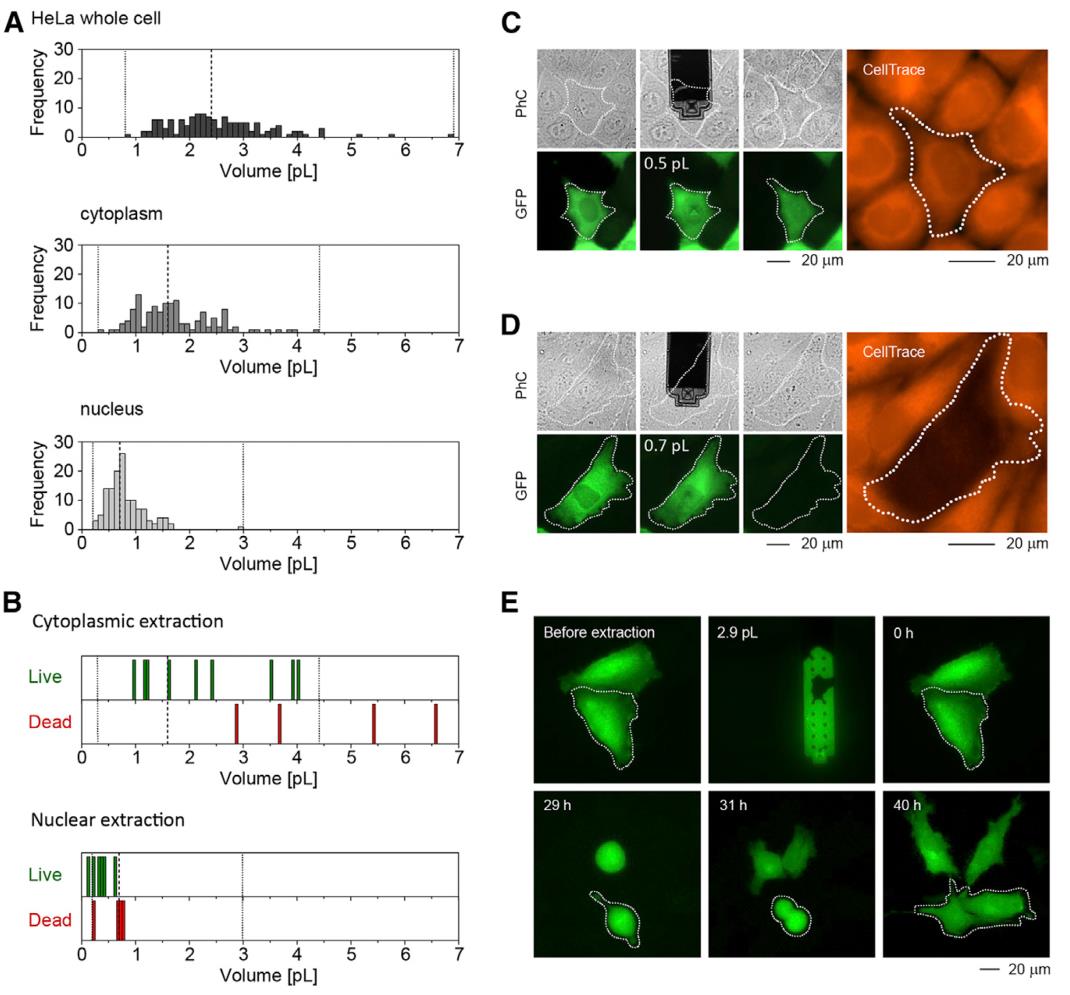
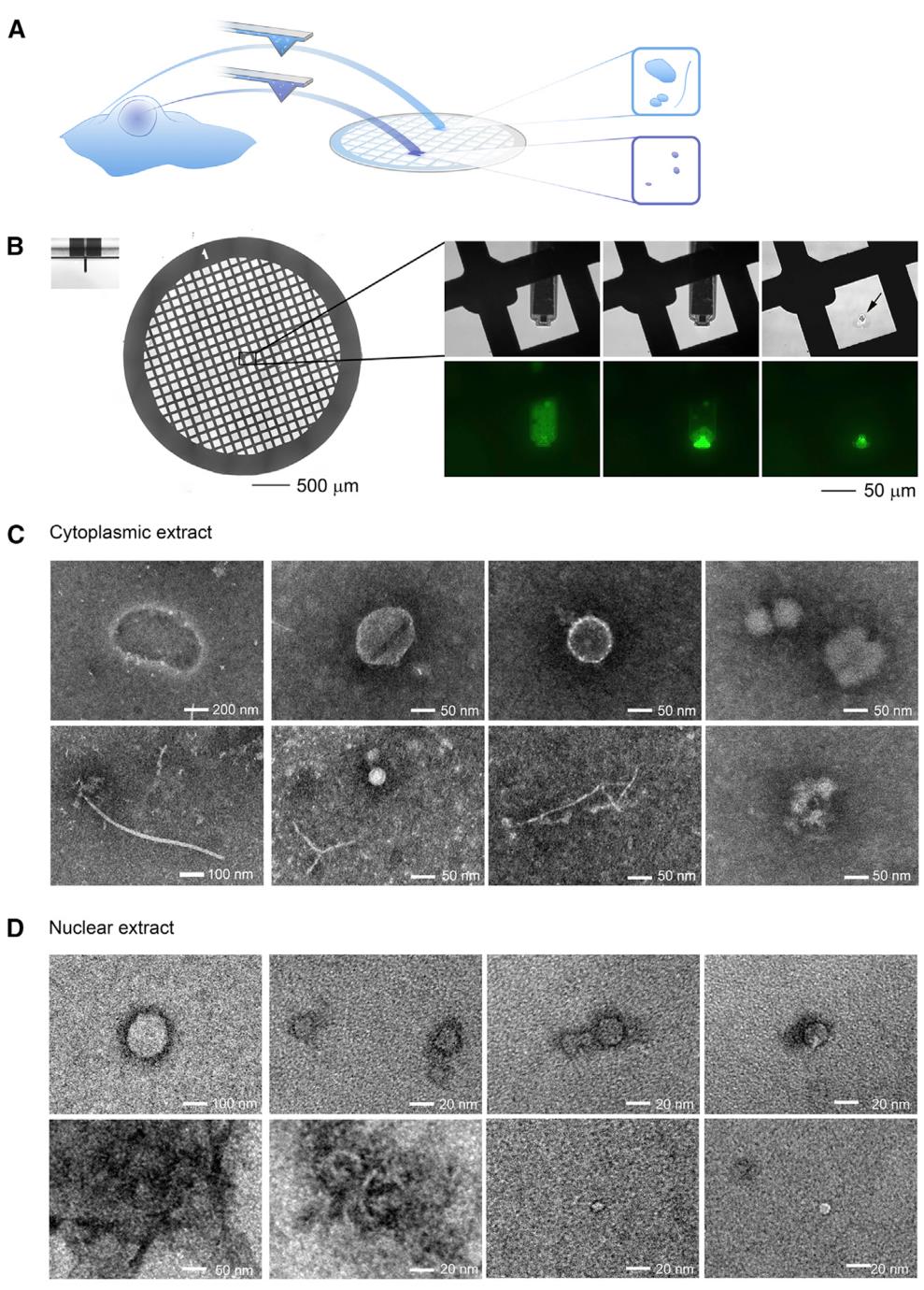
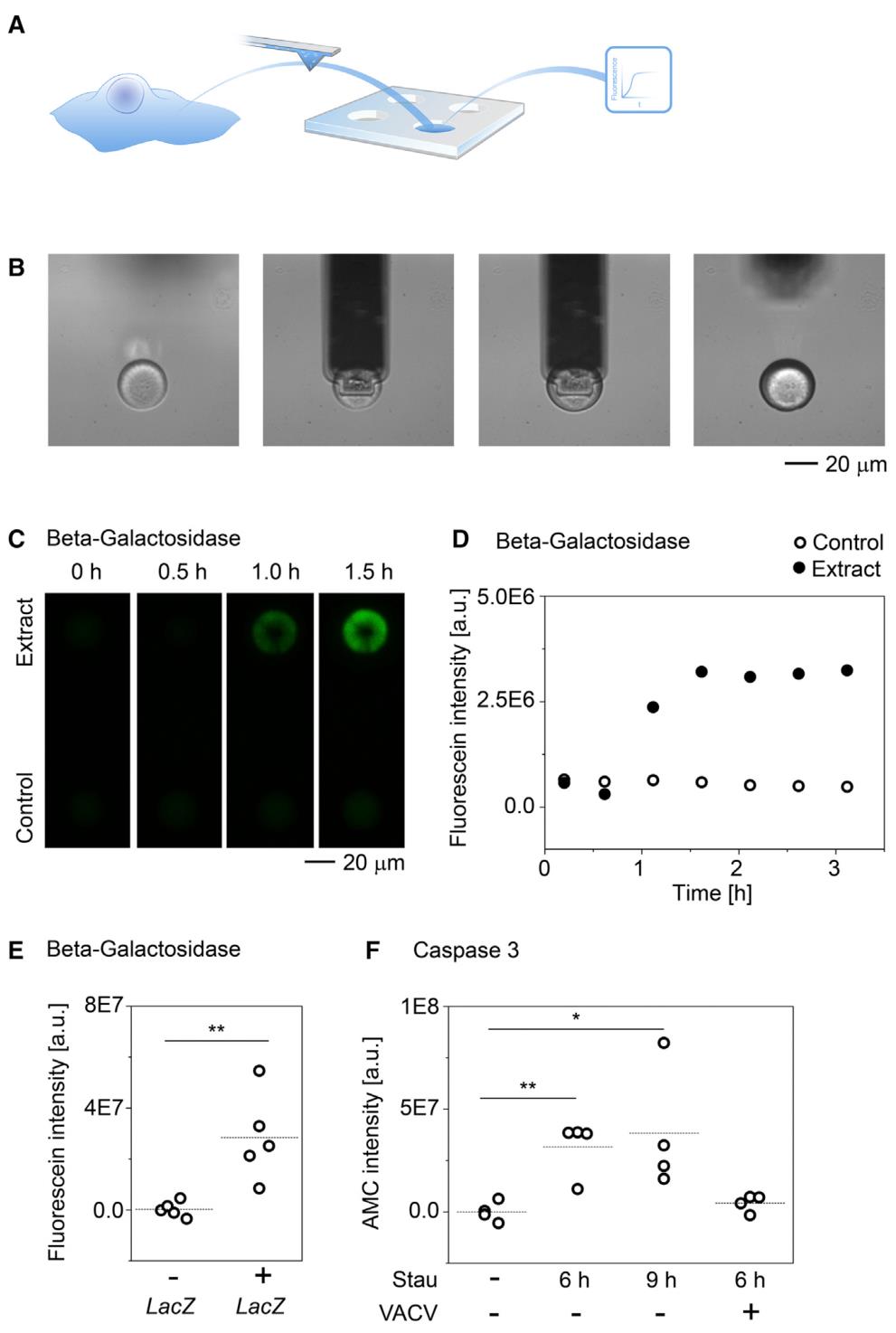
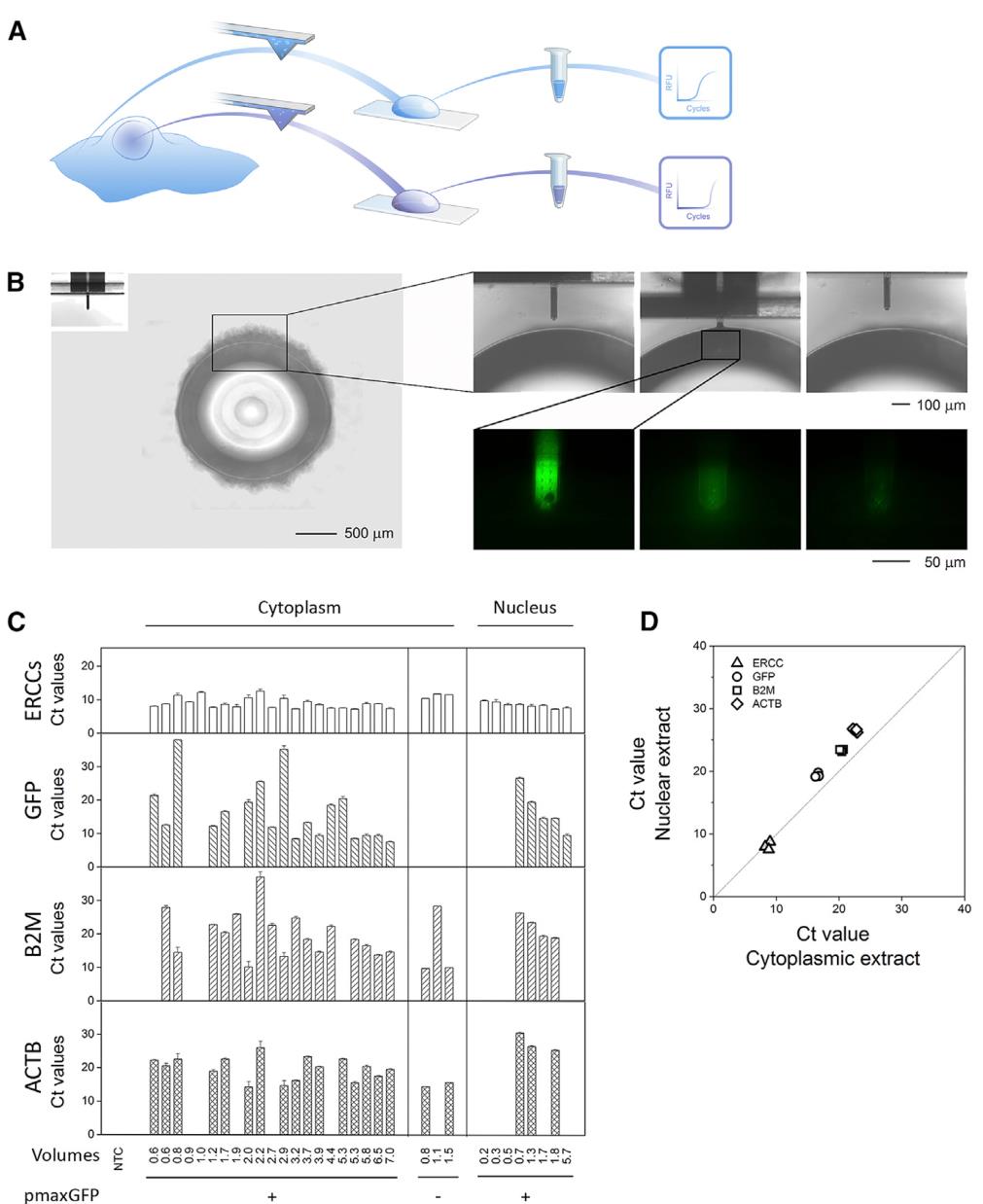
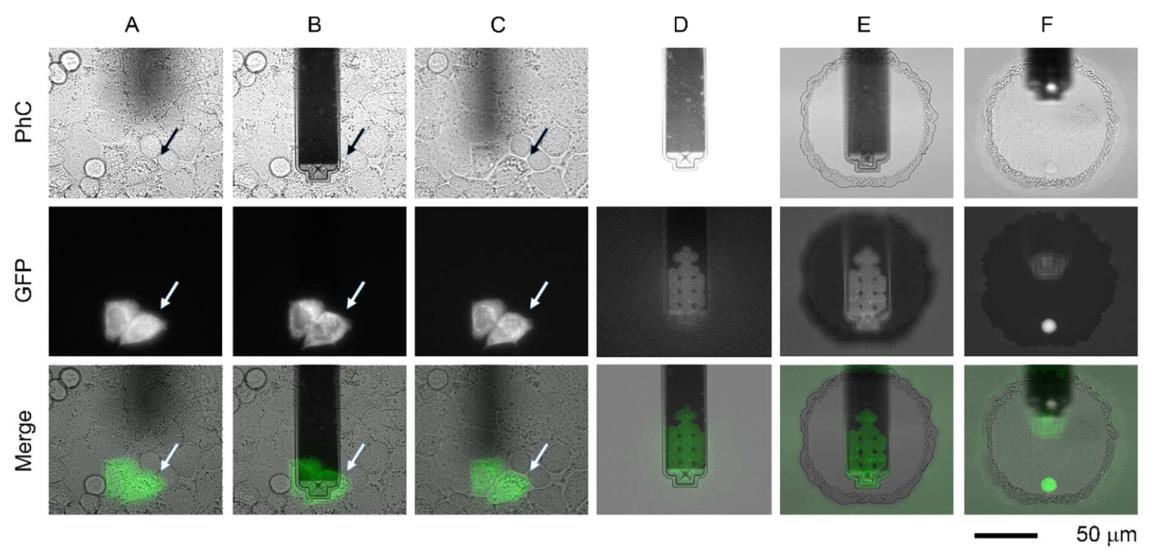
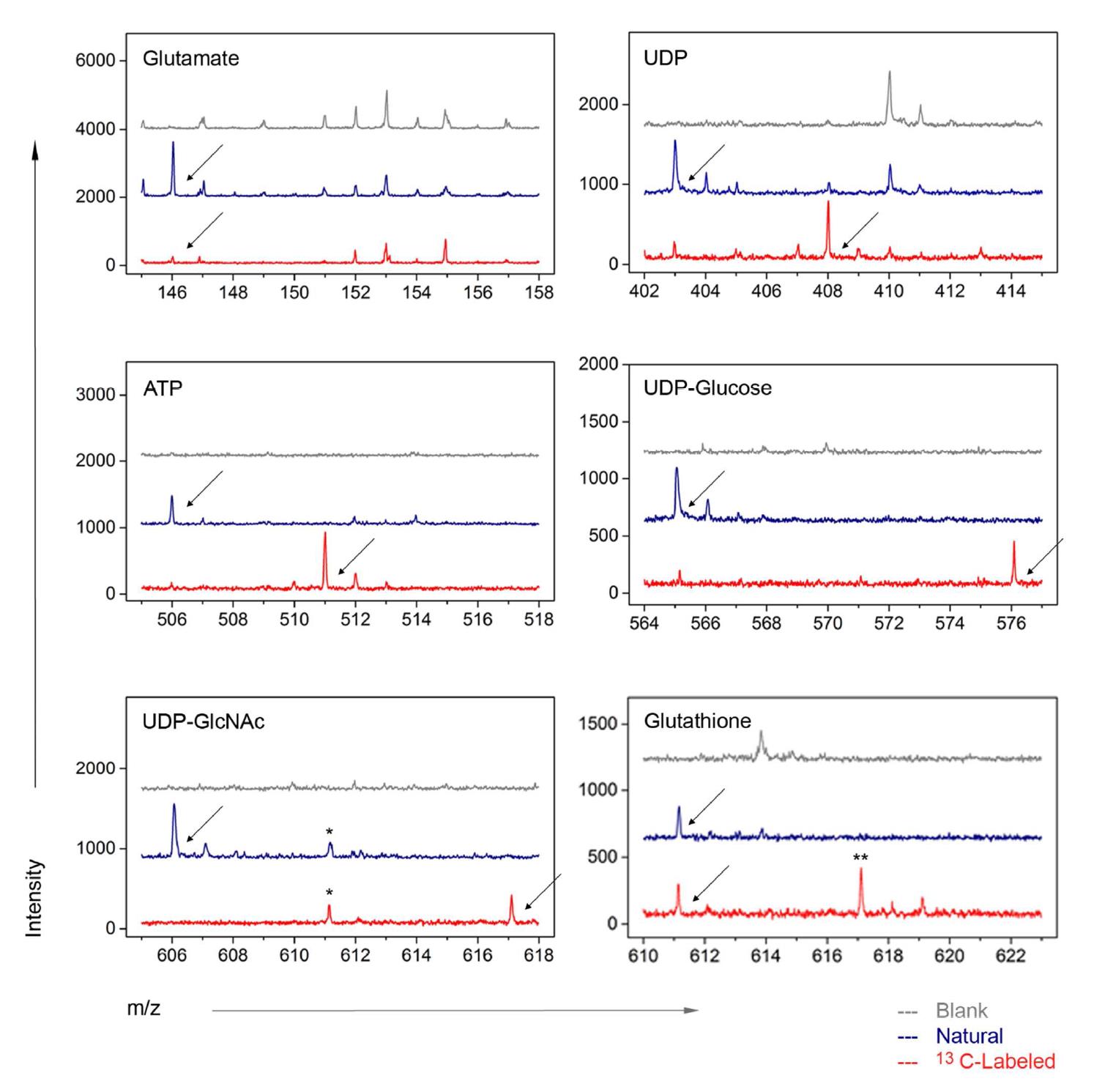
单细胞注射、提取
• W. Chen, O. Guillaume-Gentil, P. Y. Rainer, C. G. Gäbelein, W. Saelens, V. Gardeaux, A. Klaeger, R. Dainese, M. Zachara, T. Zambelli, J. A. Vorholt & B. Deplancke. Live-seq enables temporal transcriptomic recording of single cells. (2022) Nature
• Q. Hu, J. Lu, X. Zhang, R. Liu & S-H. Yang. Mitochondria transplantation/transfer between single cells. (2022). Journal of Cerebral Blood Flow & Metabolism.
• C. Gäbelein, Q. Feng, E. Sarajlic, T. Zambelli, O. Guillaume-Gentil, B. Kornmann & J. Vorholt. Mitochondria transplantation between living cells. (2022). PLoS Biology.
• O. Guillaume-Gentil, C. G. Gäbelein, S. Schmieder, V. Martinez, T. Zambelli, M. Künzler & J.A. Vorholt. Injection into and extraction from single fungal cells. (2022). Communications Biology.
• M. Li, L. Liu & T. Zambelli. FluidFM for single-cell biophysics. (2021). Nano Research. doi: 10.1007/s12274-021-3573-y FluidFM for single-cell biophysics (2021) Nano Research.
• Y. Cui, X. Lyu, L. Ding, L. Ke, D. Yang, M. Pirouz, Y. Qi, J. Ong, G. Gao, P. Du & R.I. Gregory. Global miRNA dosage control of embryonic germ layer specification. (2021) Nature.
• P. Saha, T. Duanis-Assaf & M. Reches. Fundamentals and Applications of FluidFM Technology in Single-Cell Studies. (2020) Advanced Materials Interfaces.
单细胞分选
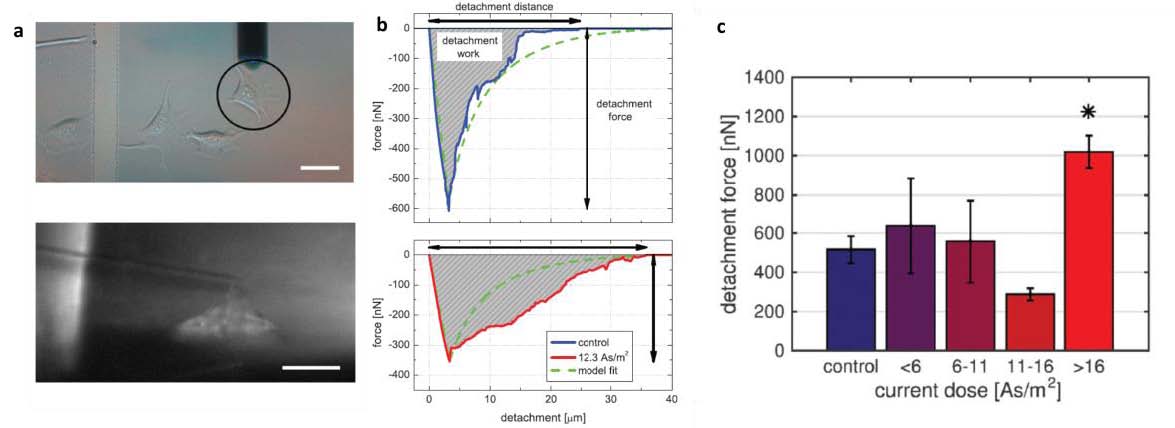
• F. Weigl, C. Blum, A. Sancho & J. Groll. Correlative Analysis of Intra- versus Extracellular Cell Detachment Events vis the Alignment of Optical Imaging and Detachment Force Quantification. (2022). Adv. Mater. Technol.
• P.W. Doll, K. Doll, A. Winkel, R. Thelen, R. Ahrens, M. Stiesch & A.E. Guber. Influence of the Available Surface Area and Cell Elasticity on Bacterial Adhesion Forces on Highly Ordered Silicon Nanopillars. (2022). ACS Omega.
• A.G. Nagy, N. Kanyó, A. Vörös, I. Székács, A. Bonyár & R. Horvath. Population distributions of single-cell adhesion parameters during the cell cycle from high-throughput robotic fluidic force microscopy. (2022). Scientific Reports.
• F. Pan, M. Liu, S. Altenried, M. Lei, J. Yang, H. Straub, W.W. Schmal, K. Maniura-Weber, O. Guillaume-Gentil & Q. Ren. Uncoupling bacterial attachment on and detachment from polydimethylsiloxane surfaces though empirical and simulation studies. (2022). Journal of Colloid and Interface Science.
• C. Chien, J. Jiang, B. Gong, T. Li & A. Gaitas. AFM Microfluidic Cantilevers as Weight Sensors for Single Cell Mass Measurements. (2022). BioRxiv.
• A. Sancho, M. B. Taskin, L. Wistlich, P. Stahlhut, K. Wittmann, A. Rossi & J. Groll. Cell Adhesion Assessment Reveals a Higher Force per Contact Area on Fibrous Structures Compared to Flat Surfaces. (2022). ACS Biomater. Sci. Eng.
• M. Li, L. Liu & T. Zambelli. FluidFM for single-cell biophysics. (2021). Nano Research. doi: 10.1007/s12274-021-3573-y FluidFM for single-cell biophysics (2021) Nano Research.
• W. Li, A. Sancho, W. Chung, Y. Vinik, J. Groll, Y. Zick, O. Medalia, A.D. Bershadsky & B. Geiger. Differential cellular responses to adhesive interactions with galectin-8- and fibronectin-coated substrates. (2021) Journal of Cell Science.
• A. Garitano-Trojaola, A. Sancho, R. Goetz, P. Eiring, S. Walz, H. Jetani, J. Gil-Pulido, M. DaVia, E. Teufel, N. Rhodes, L. Haertle, E. Arellano-Viera, R. Tibes, A. Rosenwald, L. Rasche, M. Hudecek, M. Sauer, J. Groll, H. Einsele, S. Kraus & M. Kortüm. Actin cytoskeleton deregulaltion confers midostaurin resistance in FLT-3 mutant acute myeloid leukemia. (2021) Communications Biology.
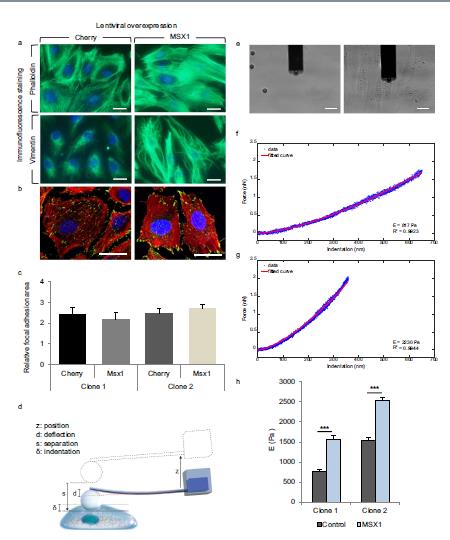
• N. Chala, S. Moimas, C. Giampietro, X. Zhang, T. Zambelli, V. Exarchos, T.Z. Nazari-Shafti, D. Poulikakos & A. Ferrari. Mechanical Fingerprint of Senescence in Endothelial Cells. (2021) Nano Letters.
• Y. Guo, F. Mei, Y. Huang, S. Ma, Y. Wei, X. Zhang, M. Xu, Y. He, B.C. Heng, L. Chen & X. Deng. Matrix stiffness modulates tip cell formation through the p-PXN-Rac1-YAP signaling axis. (2021) Bioactive Materials.
• M. Koehler, S.J.L. Petitjean, J. Yang, P. Aravamudhan, X. Somoulay, C. Lo Giudice, M. A. Poncin, A.C. Dumitru, T.S. Dermody & D. Alsteens. Reovirus directuly enganges integrin to recruit clathrin for entry into host cells. (2021) Nature communications, 12, 2149.
• M. Mathelié-Guinlet, F. Viela, J. Dehullu, S. Filimova, J.M. Rauceo, P.N. Lipke & Y.F. Dufrêne. Single-cell fluidic force microscopy reveals stress-dependent molecular interactions in yeast mating. (2021) Commun Biol.
其他
• C. Shen, Z. Zhu, D. Zhu, C. van Nisselroy, T. Zambelli & D. Momotenko. Electrochemical 3D printing of Ni-Mn and Ni-Co alloy with FluidFM. (2022). Nanotechnology.
• C. Chien, J. Jiang, B. Gong, T. Li & A. Gaitas. AFM Microfluidic Cantilevers as Weight Sensors for Single Cell Mass Measurements. (2022). BioRxiv.
• I. Demi, I. Lüchtefeld, C. Lemen, E. Dague, P. Guiraud, T. Zambelli & C. Formosa-Dague. Probing the interactions between air bubbles and (bio)interfaces at the nanoscale using FluidFM technology. (2021). Journal of Colloid and Interface Science.
• C. Müller-Renno, D. Remmel, M. Braun, K. Boonrod, G. Krczal & C. Ziegler. Producing plant virus patterns with defined 2D structure. (2021). PSS Applications and materials science.
• S. Wang, S. Liu, A. Sulkanen, J.M. Fox, X. Jia & G. Liu. Controlled Molecular Assembly of Tetrazine Derivatives on Surfaces. (2021). CCS Chem. (2021) CCS Chem.
• A. Saftics, B. Türk, A. Sulyok, N. Nagy, T. Gerecsei, I. Szekacs, S. Kurunczi & R. Horvath. Biomimetic dextran-based hydrogel layers for cell micropatterning over large areas using the FluidFM BOT technology. (2019) Langmuir, ACS.Langmuir.8b03249.
• J. Zhang, H. Yu, B. Harris, Y. Zheng, U. Celik, L. Na, R. Faller, X. Chen, D. R. Haudenschild, G. Liu. New Means to Control Molecular Assembly (2020) ACS Publications.